|
Selected Notes from a ten week course by Caspar Hewett
Introduction: Evolution by Natural Selection
evolution n. the doctrine according to which higher
forms of life have gradually arisen out of lower
natural selection evolution by the survival of the
fittest with inheritance of their fitness by the next generation
gene, n. one of the units of DNA, arranged in
linear fashion on the chromosomes, responsible for passing on specific
characteristics from parents to offspring. –adj. genic of or
relating to a gene. –ns. genome the full set of chromosomes
of an individual: the total number of genes in such a set; genotype
genetic or factorial constitution of an individual: group of individuals all of
which possess the same genetic constitution. Chambers 20th Century Dictionary
A gene is life's way of remembering how to perpetuate
itself. Genethics, Suzuki & Knudtson (1990)
A gene is any portion of chromosomal material
that potentially lasts for enough generations to serve as a unit of natural
selection. The Selfish Gene, Dawkins (1976).
DNA and RNA
Genes are the basic vehicle of biological inheritance. They
have a chemical memory which is recorded in the internal structures of a family
of biological molecules, the nucleic acids: deoxyribonucleic acid
(DNA) and ribonucleic acid (RNA).
A human cell contains 46 strands of DNA, called
chromosomes, in its nucleus, 22 matched pairs and two sex chromosomes.
This DNA contains the information necessary for synthesising proteins and for
regulating cellular processes.
DNA molecules have two distinct strands which
are held together by weak hydrogen bonds to form a double helix. Each strand is
made up of a series of small molecules called nucleotides. Only four
nucleotides are used, and they are the same in all animals and plants. They
each contain a deoxyribose sugar, a phosphate, and one of four kinds of
nitrogen-containing bases, which are usually denoted by the first
letters of their names: Adenine (A), Thymine (T), Guanine
(G) and Cytosine (C).
Each base has distinctive chemical characteristics which
allow it to bond with only one of the other three bases. Thymine will combine
only with Adenine and Cytosine will combine only with Guanine. Thus there are
four possible combinations; (T-A), (A-T), (C-G), (G-C). This is extremely
important in that it causes the two strands to be complementary to each other -
when they separate, each strand acts as a template to form its complement.
DNA encodes the genetic information for producing proteins
in the order of the bases along the DNA. The proteins themselves are made up of
amino acids. A three-base sequence along the DNA contains the instructions to
make one amino acid. RNA provides the mechanism though which DNA transmits its
message to the cytoplasm of the cell.
RNA differs from DNA in that it has ribose sugars and it has
another base, Uracil, which replaces T alongside A, G and C. Uracil
bonds with adenine forming (U-A) and (A-U) combinations. RNA is usually a
single-stranded molecule forming a single helix.
Some Definitions
A protein is any member of a group of complex
nitrogenous substances that play an important part in the bodies of plants and
animals. Proteins are compounds of carbon, hydrogen, oxygen, nitrogen, usually
sulphur, often phosphorus etc.
Amino acids are fatty acids which are the building
blocks of proteins.
A gamete is a reproductive cell. In the case of
sexual reproduction, an egg-cell or sperm-cell.
A species is ‘a group of individuals sharing
common characteristics, a group (sometimes rather arbitrarily defined) of
closely allied mutually fertile individuals showing constant differences from
allied groups, placed under a genus (biol.)’ (Chambers
20th Century Dictionary)
Speciation is the fragmentation of one species into
two or more descendant species.
Genetic drift is a chance mechanism that alters gene
frequency from generation to generation without the guiding determinism of
natural selection (due to Sewall Wright).
Taxonomy is the classification of plants and animals
into a detailed hierarchy consisting of phyla, classes, orders, families,
genera, species and sub-species or varieties.
A strategy is a term used for a pre-programmed
behavioural policy adopted by an animal, i.e. it is presumed not to be
conscious.
An Evolutionarily Stable Strategy (ESS) is a strategy
which, if most members of a population adopt it, cannot be bettered by an
alternative strategy.
Convergence is a term used to describe instances
where species from different genetic lineages develop similar physical
characteristics
A
phenotype is an observable characteristic of an organism produced by the
interaction of its genes and environment e.g. eye colour, having four legs,
inherited behavioural patterns.
A deme is a local
breeding population of a species.
An avatar is a local economic population of a species.
soma is a term for the body of an organism excluding the germ cells.

The Tree of Life
Darwin and Natural Selection
Charles Robert Darwin published On The Origin of Species by Means of
Natural Selection in 1859. He wanted to establish three things;
- The notion of evolution. That all organisms are descended from a
common ancestor.
- That life on Earth evolved by the process of natural selection.
- Life has existed for the incredible length of time necessary for the
diversity of life which exists today to have evolved in this way.
Although there were other theories of evolution around at
the time, Darwin’s great contribution was the provision of the mechanism
by which it took place. The consensus at the time was that evolution had not
taken place. Species were considered to be distinct and static rather than as
interrelated and changing. Darwin’s vision was one in which change is
inevitable and continuous. Species might seem stable but are actually
constantly evolving themselves out of existence.
Darwin’s knowledge of animal husbandry (in particular
breeding pigeons) and selective plant breeding informed his idea of natural
selection. Through selective breeding, bloodlines can be changed fairly
quickly. Darwin saw that the attributes of those individuals who were more
successful at surviving and breeding than others would spread through the
population by a similar but slower process. Those better suited to a particular
environment in any generation would have an advantage that would ensure that
the next generation contained a high percentage of their offspring. Organisms
should then become well adapted to the environment in which they live.
Darwin, of course, did not know what the vehicles of
heredity are (genes) nor its mechanics. The concept of natural selection
depended only on the basic observations that individual organisms differ, and
that they tend to resemble their parents.
Mendelian Genetics
The foundation for the field of genetics was laid by the
Augustine monk Gregor Mendel in the nineteenth century. Before he published his
work, the vehicle of inheritance was thought to be blood-borne substances which
mingled together to form offspring. Individuals were seen as being the average
of their parents. Mendel’s work proved otherwise. In 1865 he published the
results of his experiments with selective breeding using hybrid varieties of
the common garden pea, Pisum sativum.
He took advantage of two useful properties of peas:
- Each plant has both male and female organs. This means that any female
flower can be fertilised by pollen from any male. This includes the possibility
of self-fertilising plants.
- They have true-breeding lines within which every individual looks the same.
Different lines have different physical characteristics such as stem length,
seed texture and colour.
Mendel discovered, by following inheritance of a number of
selected physical characteristics, that inherited traits seemed to be
controlled by pairs of discrete hereditary ‘particles’. Adult
plants had two copies of each particle, but pollen and egg carried only one.
These ‘particles’, of course are what we describe as genes
today.
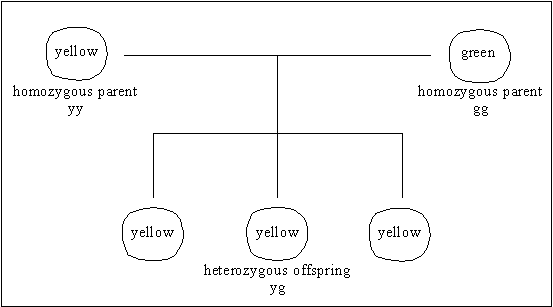
Mendelian cross showing dominance
Mendel also found that certain genes seemed to exert a more
decisive influence on inherited traits than others. These genes are described
as dominant (the gene for yellow peas, smooth seed) and those they
dominate are described as recessive (green peas, wrinkled seed). Since
each pea plant has two complete sets of genes, each trait could be determined
by either of two possible alternative genes, or alleles. The plant is
described as homozygous if the alleles are the same and
heterozygous if they are different.
Mutation and Discontinuity
Mendel's work was largely undiscovered until the
beginning of the twentieth century, and as such was completely unknown to
Darwin. However, with the rediscovery of Mendel’s work in the early part
of the century came a decline in the perceived importance of natural selection.
This was reinforced by the discovery of mutation by DeVries. An alternative
theory of evolution which emphasised the importance of mutation was proposed
and was thought by many to supersede the theory of evolution by natural
selection. Early experiments in genetics showed mutations having large-scale
effects, which seemed to confirm the existence of rapid, discontinuous
evolutionary change. However this had to be tempered with the observation that
large mutations were often harmful and sometimes lethal.
There are echoes of this idea in the work of the geneticist
Richard Goldschmidt in the 1930s and 1940s. He observed significant
discontinuity when studying the genetic variation of the silkworm moth. He
attempted to explain this by invoking large-scale mutations or "hopeful
monsters". Goldschmidt and the other saltationists were ridiculed
by the Neo-Darwinists for their ideas and provided a focus around which the
Neo-Darwinists united.
Theodosius Dobzhansky, also writing in the 1930s stressed
the importance of geographic and reproductive isolation, another aspect of
discontinuity. Dobzhansky argued that when a species divides into two or
more discrete reproductive communities natural selection will tend to hone the
adaptations of each community to their particular environment, which would tend
to minimise variation within new species. Dobzhansky went even further in
saying that isolation between species is a direct outcome of the evolutionary
process.
Neo-Darwinism – The Modern Synthesis
The 1920s saw the emergence of Neo-Darwinism, which was
founded by Ronald Fisher, J. B. S. Haldane and Sewall Wright. They reconciled
the idea of evolution by natural selection with the discontinuous, particulate
nature of genes. This was the essence the modern synthesis of Darwin's theory
and Mendelian genetics.
The new synthesis continued to develop in the 1940s, notably
with Julian Huxley’s, Evolution: The Modern Synthesis (1942) and
Bernhard Rensch’s, Evolution Above the Species Level (1947).
Natural selection was seen as the dominant force shaping evolutionary change.
Rensch expressed the view that nothing in biological nature suggests that any
evolutionary processes other than natural selection work on the natural
genetics of variation within populations.
Modern Neo-Darwinism – The Selfish
Gene
Richard Dawkins epitomises what is described by some as the
ultra-Darwinian view. In his 1976 book The Selfish Gene he presents the
theory of evolution by natural selection in a very general way as an
algorithm. First he introduces the concept of a replicator, which
is anything which is capable of making replicas of itself. He argues that the
two conditions necessary for the evolutionary process to begin are the
existence of a replicator and the possibility of small errors being made in the
copying process. Once it exists a replicator can generate copies of itself
indefinitely. Mistakes in the copying process will lead to a situation where
there is more than one variety of replicator in the population. Some new
varieties will be less effective at self-replicating than the old, some will
not be able to replicate themselves at all, but some new varieties will be
better than the old - This may be because they produce copies of themselves
more quickly than their predecessors (fecundity), because they survive
for longer and thus are able to self-replicate for longer (longevity),
or because they are less prone to errors in the copying process
(accuracy). Whatever the reason, such a replicator's descendants will
naturally come to dominate the population. The ‘fittest’ replicator
then can be thought of as the one of which there are the most surviving copies.
As the population of replicators grows there will be
increasing competition between them. This will lead over time to the evolution
of increasingly elaborate ways of surviving and replicating. Certain
characteristics will be selected for, i.e. replicators which have them
will be more abundant than those that do not. The success of a replicator will
depend on its environment, and especially on the effects other replicators have
on it and on its environment. Mutually compatible replicators will evolve and
perhaps larger vehicles which house the replicators will follow. This
does indeed appear to have been the case on earth.
The formation of the first replicating molecule was an
‘exceedingly improbable’ accident. However, once such a molecule
existed, the process above - evolution by natural selection - began. At
some point in evolutionary history mutually compatible replicators created
vehicles - cells and, later, many-celled bodies. The replicators of today are
DNA and we - that is humans, animals and plants - are their survival machines!
Dawkins thus invites us to think of the body as a colony of
genes. He emphasises that the basic unit of natural selection is best regarded
as the gene.
It is worth noting that we often use purposive language when
we speak about natural selection, for example ‘the robin evolved wings for
flying’ does not imply any purpose in the sense that someone designed the
wings with a purpose in mind - It is accepted that natural selection is a blind
process, but it is wise to be wary of using this type of language, especially
when discussing animal behaviour.
Animal Behaviour -
Altruism
Dawkins tells us to expect ‘ruthless selfishness’
in a successful gene. However, he spends much of The Selfish Gene
discussing altruistic behaviour and how it evolved. His argument hinges on the
idea that the selfish gene is not a specific piece of DNA, but is rather
‘all replicas of a particular bit of DNA, distributed throughout
the world.’ This emphasis on the distributed nature of the gene leads to
the observation that, if an animal has some way of knowing that copies of its
own genes reside in other individuals, then its genes have good reason to
programme that animal to behave altruistically towards those individuals. Often
what appears to be altruistic behaviour is actually gene selfishness. Thus
‘there are special circumstances in which a gene can achieve its own
selfish goals best by fostering a limited form of altruism at the level of
individual animals.’ This is the argument behind kin selection and,
less obviously is applied to a range of seemingly altruistic behaviour that
depends on mutual benefit of the individuals involved, both within species and
across boundaries between species.
A simple replicator
Conflicts of interest in the family
There is a real conflict of interest between parent and child. In practice,
some compromise is usually reached between the ideal situations of the parent
and its offspring.
Genes in the bodies of children are selected for their ability to outsmart
their parents, and genes in the bodies of parents are selected for their
ability to outsmart their offspring.
Resources invested by parents on offspring
- Food
- Energy and time expended obtaining food and maintaining the nest or home
- Time spent teaching children
- Risks taken to protect young
Definition: Parental Investment (PI) is any
investment by a parent in one of her (his) offspring that increases the chance
that the offspring will survive at the expense of that parent’s ability to
invest in any other offspring (alive or yet to be born).
Sibling Rivalry
Dawkins states that much of child behaviour is characterised
by selfish greed. However, in the case of monogamous species, the relation of
an individual child to its siblings is the same as that of its mother, ½.
That is, on average, the child will share half of its genes with any one of its
siblings. Thus we would expect the child to want some of the parental
investment to go to its brothers and sisters, especially if they are
contemporaries. Theory would predict that the child will stop grabbing extra at
that point where the resulting cost to its sibling is twice the benefit to
itself.
Favouritism
There is no genetic reason for a mother to have favourites.
She has the same relatedness to all of her children – they each have half
of her genes. Thus her optimal strategy is to have as many children as possible
that she can successfully raise to survive to reproductive age.
Weaning
Weaning is another example where there is a conflict of
interest between mother and child. The disagreement is essentially one over
timing. We would expect the mother to want to wean child when she reaches a
point when her PI could be better spent on having another child, or even on
other relatives such as grandchildren, nieces, nephews. In contrast, the child
will not want to be weaned until the cost to its unborn sibling is twice the
benefit to itself.
Conflict between mates
In the past sexual behaviour was sometimes considered as a co-operative
venture undertaken for the good of the species or at least for mutual benefit
of the two partners. In contrast, from the selfish gene perspective (following
the work of R.L. Trivers), the relationship of sexual partners is seen as one
based on mutual mistrust and exploitation!
There is a fundamental asymmetry between the sexes in most species. Male
gametes (sperm) are more mobile, smaller and more numerous than those of
females (eggs). This means that a female can have only a limited number of
offspring, whereas a male can have a virtually unlimited number, provided that
he can find females willing to mate with him.
In some fungi and other primitive organisms there are not two distinct
sexes, this is known as isogamy. Rather than having two types of sex
cells, sperms and eggs, all of the gametes are the same –
isogametes. A new individual is formed by the fusion of two isogametes,
each produced by meiotic division.
There are a variety of reasons why the asymmetry between the sexes may have
evolved from an isogamous state (See exercises).
Desertion
The asymmetry discussed above puts the female at a
disadvantage. She invests much more than the male initially and thus is open to
desertion by her mate. If there is any chance that the female will successfully
rear their young without his help the male’s best strategy is to leave and
find another mate. Thus we would expect to find a bias towards lack of paternal
care to evolve, and it is observed in some species. However there are many
other selection pressures which could act against this tendency.
Desertion strategies
Given that the male has deserted, the female can adopt one
of a number of strategies:
Deceive another male – There would then be
selection pressure against gullibility in males. This can be one reason why a
prolonged period of courtship can benefit a male – so that he can be sure
that any children she produces are his.
She can abort - This is likely to be a preferred option if the child
has only just been conceived. If she is not likely to deceive another male this
strategy could be to mutual advantage.
Rear the young herself - which is most likely to pay if they are quite old.
Counter-desert - It could pay either partner to be the first to desert!
Courtship
In considering the possibility of being deserted the female
can play her ace – she can refuse to copulate. Dawkins proposes that, in
choosing a mate, a female has two main strategies open to her.
Domestic-bliss strategy : The female looks for
males who exhibit signs of fidelity and domesticity. One way she can achieve
this is by being coy – the male has to prove his faithfulness and
perseverance. Courtship rituals can represent some considerable investment by
the male before copulation. In the case of some birds the male has to build a
nest before mating, which then represents a direct investment by the male in
the eggs! Breeding with last season’s mate also a good strategy for a
female. He-man strategy : The female goes for the best genes, accepting that she
will get no help from the father. She refuses to copulate with all but the best
males. In this situation only the best males would reproduce. The female would
look for evidence of ability to survive and reproduce – qualities such as
strong muscles, long legs, age. It is worth noting that once females begin to
select their mates on the basis of a useful attribute such as long legs, then
we would expect natural selection to favour that attribute purely because it is
attractive to females.
Note:
- Previous investment by either parent in their offspring is no deterrent to
desertion provided there is a good chance that those offspring will survive
without them!
- Behavioural strategies such as the domestic-bliss strategy often depend on
recognition between individuals and thus is one of many reasons why many
animals recognise other individuals.
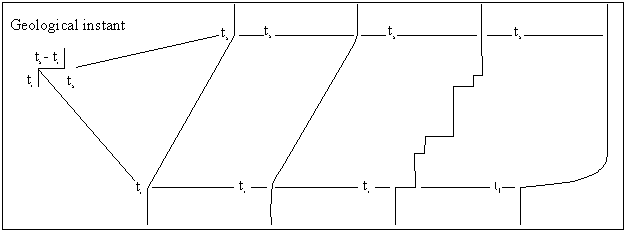
Stasis
The notion of stasis is based on the observation that
species appear to change very little throughout their existence. Darwin
acknowledged the abrupt appearance of species in the fossil record and their
apparent stability in the sixth edition of On The Origin of Species.
Darwin had his own reasons for emphasising gradualism in the evolutionary
process – he was trying to establish the notion of evolution itself! The
existence of stasis is seen by those in the ‘naturalist’ camp,
notably Stephen Jay Gould and Niles Eldredge, as requiring explanation; there
is no doubt that natural selection is the motor by which adaptive change takes
place but, they argue, the question remains why adaptive change takes place
when it does.
Habitat Tracking
Given an environmental change there are three possible
outcomes for a species:
- A geographical move
- Adaptation to the new environment.
- Extinction
The notion of habitat tracking is simply based on the
observation that when environmental change takes place, it is common for
species to move geographically – finding a suitable habitat elsewhere.
According to Niles Eldredge, adaptation is the least likely of the three
outcomes above, with extinction in second place!
Punctuated Equilibria
Niles Eldredge describes punctuated equilibrium as
"a molding, in essence, of the pattern of stasis with the recognition that
most evolutionary change seems bound up with the origin of new species - the
process of speciation." The importance of the theory is that it tells us
something about the context of evolutionary change - why evolutionary
change happens when it does. He and Stephen Jay Gould introduced the theory in
their 1972 paper Punctuated equilibria: an alternative to phyletic
gradualism in an attempt to deal with some of the problems for evolutionary
theory highlighted by palaeontology:
- Persistent patterns of little or no change are found within fossil samples
in a particular location (stasis)
- Evolutionary novelty usually appears abruptly in the fossil record and it
is very difficult to know whether an organism evolved at the location where its
fossil was found or whether it came from elsewhere.
- Only a tiny percentage of all organisms ever appear in the fossil record!
Eldredge and Gould were inspired by the work of Dobzhansky
and Mayr on species and the speciation process, especially the idea that
descendant species derive from their ancestral species through geographic
isolation. Paterson's Specific Mate Recognition System provides further
justification for the idea.
Eldredge and Gould begin their 1972 paper by noting how
difficult it is to develop new theory when our interpretation of facts depends
heavily on old theory: ‘New pictures must cast their influence before
facts can be seen in different perspective.’ They state that the picture
of ‘phyletic gradualism’ has dominated palaeontology and that,
because of this, all breaks in the fossil record are treated as imperfections
in the record rather as real phenomena requiring explanation. They identify the
following as the tenets of phyletic gradualism:
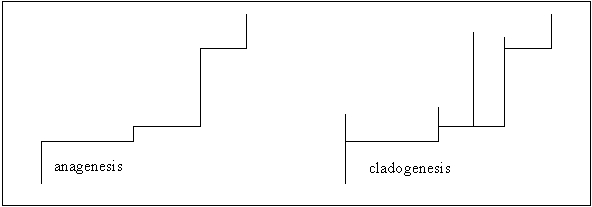
New species arise by the transformation
of an ancestral population into its modified descendants.
- The transformation is even and slow.
- The transformation involves large numbers, usually the entire ancestral
population.
- The transformation occurs over all or a part of the ancestral species’
geographic ranges.
Eldredge and Gould argue that, if evolution does indeed
occur in this gradualist fashion, then the fossil record would be expected to
‘consist of a long sequence of continuous, insensibly graded intermediate
forms linking ancestor and descendant.’ In practice there are usually
morphological breaks found in postulated phyletic sequences, but these are
traditionally put down to the imperfections of the fossil record. This approach
‘renders the picture of phyletic gradualism virtually unfalsifiable.’
Further, even the term "morphological break" presupposes the
continuous nature of such sequences. Thus, they argue that only an alternative
picture is capable of shedding light on cases in which breaks are found and to
treat them as real phenomena.
They advance the theory of allopatric (or
geographic) speciation as providing an alternative picture. This
is a theory that developed through the study of modern species’
distribution, ecology and behaviour. According to this theory ‘new species
can arise only when a small local population becomes isolated at the margin of
the geographic range of its parent species.’ These peripheral
isolates evolve into a new species if isolating mechanisms develop
which make reproduction impossible with the parent species should their paths
cross at a later date. This means that we would not expect new fossil species
to appear in the same place as their ancestors! Speciation relies on the
limited gene pool of the small initial population to speed up morphological
change – large gene pools tend to be conservative. This suggests a picture
of (relatively) rapid change initially followed by stability once the new
species is established. Thus ‘it is likely that the two species will
display their greatest difference when the descendant species first
appears.’ This pattern is borne out by the fossil record – which can
thus be taken as much more reliable than tradition dictates.
Eldredge and Gould see the picture of phyletic gradualism as
inadequate to explain speciation because it ‘fails to recognize that
speciation is primarily an ecological and geographic process.’ The theory
of punctuated equilibria considers speciation to be a component of the
evolutionary process: ‘The history of evolution is not one of stately
unfolding, but a story of homeostatic equilibria, disturbed only
"rarely" (i.e., rather often in the fullness of time) by rapid and
episodic events of speciation.’ Eldredge describes punctuated equilibria
as providing ‘the context - the boundary conditions - for much of what
really matters in terms of adaptive change in evolutionary history.’ Thus
speciation is the key to understanding why we find long periods of stasis being
interrupted by shorter periods of (relatively) rapid evolutionary change.
Eldredge and Gould found themselves accused of proposing a
form of saltationism. To explain why this is not the case Eldredge draws
attention to the time-scales relevant to palaeontology:
- Most species of marine vertebrates last between 5 million and 10 million
years.
- Terrestrial animals do not tend to last quite as long, mainly thanks to the
more frequent environmental changes experienced on land.
- The fossil record only has a resolution of tens of thousands of years.
- Eldredge estimates the time required for speciation to be five to fifty
thousand years.
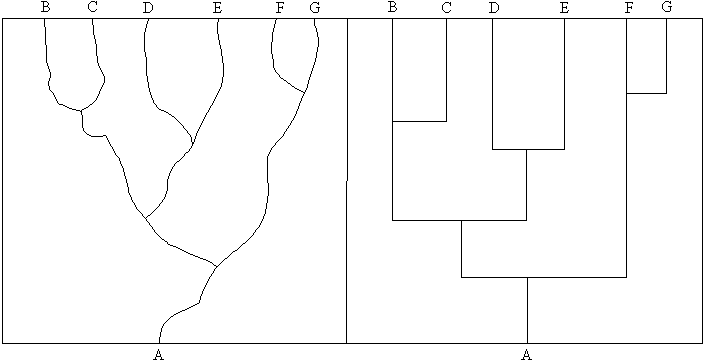
In Darwin's Dangerous Idea (1995) Daniel Dennett
questions whether there is anything new in the punctuated equilibrium thesis.
Like Eldredge, he draws attention to the problem of time-scale. The left hand
diagram in Figure 1 represents the orthodox Neo-Darwinian view, while the
right hand diagram represents punctuated
equilibria. The horizontal axis represents changes in design and the vertical
axis is time. On close inspection – when the time-scales are not quite so
vast – Dennett questions whether there is any real difference between
these two views, especially when Gould has clearly stated that he does not
propose saltations – speciation takes place gradually – the
apparently horizontal sections of the right hand diagram are actually a ramp
over the speciation period.
|